Thème de recherche n°2 : Utilisation des sucres aminés par E. coli et B. subtilis.
Les bactéries sont capables d'utiliser une grande variété de substances en tant que sources de carbone et d'énergie. Les sucres aminés sont une bonne source d'azote et de carbone. Ils sont, de plus, des éléments essentiels car ils forment le squelette de peptidoglycane de la paroi cellulaire chez les bactéries. Nos études s’intéressent à la régulation des gènes responsables de l'utilisation des sucres aminés, la N-acétylglucosamine (GlcNAc) et la glucosamine (GlcN), ainsi que les rôles physiologiques de ces gènes et leur influence sur le recyclage du peptidoglycane chez E. coli.
Comparaison de la régulation par les facteurs de transcription homologues, Mlc et NagC chez Escherichia coli.
Chez E. coli et B. subtilis les sucres aminés sont assimilés par le système phosphotransférase (PTS), une cascade de phosphorylation complexe, qui se traduit par le transport et la phosphorylation simultanée des sucres. Chez E. coli, les gènes pour l'utilisation de la GlcNAc sont contrôlés par le répresseur NagC (Fig. 1).
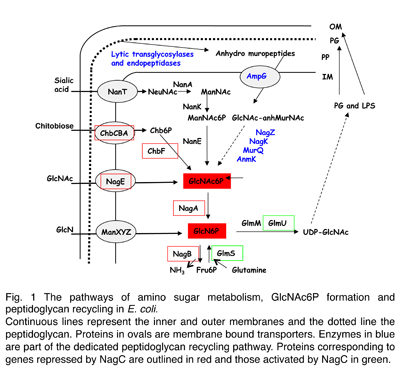
Mlc, un orthologue de NagC chez E. coli, contrôle les gènes de l'absorption du glucose par le PTS. Ces deux facteurs de transcription sont 70 % similaires et ils reconnaissent des séquences opératrices très semblables. Nous avons montré que la spécificité de la fixation à l’opérateur est déterminée par une séquence dite "linker", qui forme un aile ("wing") au motif hélice-coude-hélice de fixation à l'ADN et que les mutations dans cette séquence permettent de basculer la régulation de NagC vers Mlc et vice versa. En revanche, les signaux d'induction pour les deux protéines sont très différents. Pour NagC l’inducteur est le GlcNAc-6P produit lors du transport de la GlcNAc par le PTS. Quant à Mlc, elle se lie aux membranes qui contiennent le transporteur du glucose déphosphorylé. Mlc est donc sensible à l'activité de transport du glucose. Nous avons identifié les acides aminés importants pour ces deux mécanismes de signalisation. L’homologue de Mlc chez V. cholerae montre, au moins chez E. coli, un mécanisme d'induction hybride. Il est séquestré aux membranes pendant le transport de la GlcNAc, non pas du glucose, et de plus une fraction de l'induction peut être attitrée au GlcNAcP.
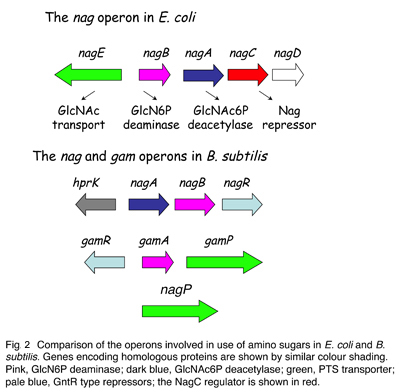
Les enzymes requises pour l'utilisation de la GlcNAc sont conservées chez B. subtilis mais elles sont soumises à une régulation différente. Contrairement à E. coli, et à la plupart des autres Bacilli, B. subtilis pousse mieux avec GlcN qu'avec GlcNAc en tant que source de carbone. Nous avons montré que cela est dû à la présence d'un opéron unique (gamAP) codant pour des gènes dupliqués pour le transport et le métabolisme de la GlcN (Fig. 2). Les gènes nag et gam de B. subtilis sont régulés par les facteurs de transcription homologues appartenant à la famille GntR (qui n'ont pas de rapport avec NagC) et leurs propriétés de liaison à l'ADN et de reconnaissance de l'inducteur ont fortement divergé.
Les gènes homologues, codants les désaminases du GlcN6P (nagB) se retrouvent chez un grand nombre d’organismes. Chez certaines espèces (e.g. E. coli et les mammifères) les enzymes sont allostériques alors qu’elles ne le sont pas chez d'autres espèces (B. subtilis et les levures). En collaboration avec Mario Calcagno (UNAM, Mexico), nous étudions le rôle et les conséquences de la régulation allostérique chez E. coli et celui de l'absence d’une telle régulation chez B. subtilis, en utilisant plusieurs techniques telles que la génétique, la biochimie, la biophysique et la métabolomique.